Se også: ketogen diæt; diabetisk ketoacidose.
Almindelighed
Tidligere troede man, at ketonlegemer skyldtes overdreven stofskifte forårsaget af indtagelse af for meget fedt eller diabetes.Ketonkroppe produceres derimod naturligt af vores krop: hjernen tilpasser sig til at bruge disse metabolitter under forhold af langvarig faste (hos diabetikere erstatter ketonlegemer glukosemetabolismen) Desuden kan der være en forfærdelse af ketonlegemernes vej i tilfælde af dårlig ernæring.
Hvad er ketonlegemerne
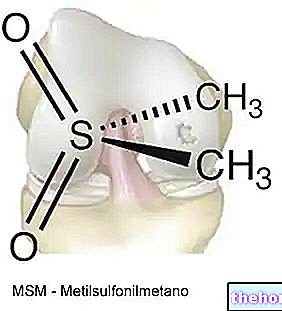
Ketonlegemer er derivater af lipider (de stammer fra lipids metabolisme, næsten udelukkende hepatisk), men har egenskaber, der får dem til at ligne sukkerarter:
- Høj inputhastighed;
- Hurtig at bruge.
Selv nogle aminosyrer, især metaboliske tilstande, kan stamme fra ketonlegemer (Leucine, Lysine, Phenylalanine, Isoleucine, Tryptophan og Tyrosine).
Biologisk rolle
- Ketonlegemer er små i størrelse, derfor transporteres de meget hurtigt (meget mere end fedtsyrer, der på den anden side har brug for transportproteiner som albumin);
- ketonlegemerne bruges næsten udelukkende af musklerne og perifere væv, men også af hjertet (20-30% af den energi, den bruger, kommer fra ketonlegemerne) og af hjernen (i tilfælde af langvarig faste).
Syntese
Ketonlegemer syntetiseres af acetylcoenzym A, som stammer fra metabolismen af fedtsyrer.
Enzymet, der katalyserer det første trin, er Β-ketothiolase, som udnytter svovl af acetylcoenzym A til at producere et Β-ketoacyl-coenzym A (det er den modsatte reaktion til den, der ses ved Β-oxidationen af fedtsyrer); denne reaktion er ikke spontan, men drives af den efterfølgende reaktion , katalyseret fra "hydroxymethylglutaryl -coenzym A -syntase og som involverer vedhæftning af et andet acetylcoenzym A, opnåelse af 3-hydroxy-3-methylglutaryl-coenzym A.
Efterfølgende griber et lytisk enzym ind, som omdanner 3-hydroxy 3-methylglutaryl-coenzym A til eddike-acetat, som er et ketonlegeme. Eddikeacetatet kan sendes til det perifere væv eller ved enzymets virkning hydroxybutyrat dehydrogenase, omdannet til 3-Β-hydroxybutyrat. Hvis eddikeacetatet er i meget høj koncentration, kan det også spontant decarboxylere til acetone.
Acetone, eddikeacetat og 3-Β-hydroxybutyrat er de tre ketonlegemer, som vi betragter; acetone er et affaldsprodukt, der produceres tilfældigt i ketonlegemernes vej og udvises ved udånding og transpiration.
Anvendelse i perifert væv
Ketonlegemer, der produceres i leveren, sendes til perifert væv.
Lad os nu se, hvad der sker, når eddikeacetat og 3-Β-hydroxybutyrat når det perifere væv. Eddikeacetatet er en Β-ketosyre, derfor kan det, hvis det aktiveres, bruges i Β-oxidationsprocessen til fremstilling acetylcoenzym A: Derfor er det nødvendigt at omdanne en Β-ketosyre til et Β-ketoacyl-coenzym A.
Når eddikeacetatet ankommer til mitokondrien i en celle i et perifert væv, udsættes det for enzymets virkning succinylcoenzym A -transferase: gennem dette enzym reagerer eddikeacetat med succinylcoenzym A (kommer fra krebs -cyklussen), og succinat og eddike -acetylcoenzym A opnås.
Ved at udnytte succinylcoenzym A, for at aktivere eddikeacetatet, hopper vi ind i krebs -cyklussen, det trin, der producerer en GTP: dette er processen, hvad angår energi, som cellen er villig til at betale for at opnå acetyleddike -coenzym A ; sidstnævnte går derefter under handlingen af Β-keto-thiolase (Β-oxidationsenzym) til at producere to molekyler acetylcoenzym A, der sendes til krebs-cyklussen.
Hvis 3-Β-hydroxybutyrat sendes til perifert væv, omdannes sidstnævnte inde i mitokondrionen til eddike-acetone ved virkningen af Β-hydroxybutyrat-dehydrogenase-enzymet med produktion af et NADH, der svarer til ca. 2,5 ATP; eddikeacetat produceret følger den tidligere beskrevne vej.
Cellen i et perifert væv trækker mere energi fra 3-Β-hydroxybutyrat frem for fra eddikeacetat, men levering af det ene eller det andet til det perifere væv afhænger af leverens energitilgængelighed.
C "er en ubetydelig mængde metaboliserede fedtsyrer, der er indeholdt i peroxisomer og ikke i mitokondrier; peroxisomer er organeller mindre end mitokondrier og rige på metalioner og peroxidaseenzymer. Peroxidaseenzymer bruger hydrogenperoxid til at fremme redoxprocesser, derfor findes der i peroxisomer er et enzymatisk system, der er i stand til at producere hydrogenperoxid.
I Β-oxidationen i peroxisomer opnås "acylcoenzym A, ved virkningen af"acylcoenzym A -oxidase (I mitokondrierne virkede derimod enzymet acylcoenzym A dehydrogenase). Også i dette tilfælde dannes trans 2,3 enoylcoenzym A, som gennemgår virkningen af et bifunktionelt enzym (det udfører samme funktion som i mitokondrier af "enoylcoenzym A hydratase og L-Β-hydroxyacylcoenzym A dehydrogenase) og omdannes således til Β-ketocylcoenzym A. Dette sidste, som i mitokondrier, undergår virkningen af Β-ketotiolase og acetylcoenzym A og et acylcoenzym A opnås med et kulstofholdigt skelet reduceret med to enheder i forhold til det startende, som vender tilbage til cirkulationen.
.jpg)













-cos-cause-e-cura.jpg)