For at kunne tale på en forståelig måde om "hæmoglobin (Hb), det er nyttigt at tage sig af det første myoglobin (Mb) som ligner meget hæmoglobin, men er meget enklere. Mellem hæmoglobin og myoglobin er der tætte slægtskabsforhold: begge er konjugerede proteiner og deres protetiske gruppe (ikke-proteindel) er gruppen heme.
Myoglobin er et globulært protein, der består af en enkelt kæde på omkring hundrede og halvtreds aminosyrer (det afhænger af organismen), og dets molekylvægt er omkring 18 Kd.
Som nævnt er den udstyret med en hæm-gruppe, der indsættes i en hydrofob (eller lipofil) del af proteinet, der består af folder, der kan tilskrives α-helix-strukturer af fibrøse proteiner.
Myoglobin består hovedsageligt af segmenter af a-helixer, der er til stede i antal otte og består næsten udelukkende af upolære rester (leucin, valin, methionin og phenylalanin), mens polære rester er praktisk talt fraværende (asparaginsyre, glutaminsyre, lysin og arginin); de eneste polare rester er to histidiner, som spiller en grundlæggende rolle i vedhæftningen af ilt til hæmgruppen.
Hæm -gruppen er en kromoforgruppe (absorberes i det synlige) og er den funktionelle gruppe af myoglobin.
Se også: glyceret hæmoglobin - hæmoglobin i urinen
Lidt kemi

Bindingen mellem protoporphyrin og jern er en typisk binding af koordineringsforbindelser, som er kemiske forbindelser, hvor et centralt atom (eller ion) danner bindinger med andre kemiske arter i et tal større end dets oxidationsnummer (elektrisk ladning). I tilfælde af hæm er disse obligationer reversible og svage.
Jernets koordinationsnummer (antal koordinationsbindinger) er seks: der kan være seks molekyler omkring jernet, der deler bindingselektronerne.
For at danne en koordinationsforbindelse kræver det to orbitaler med korrekt orientering: den ene er i stand til at "erhverve" elektroner og den anden er i stand til at donere dem.
I hæm danner jern fire plane bindinger med de fire nitrogenatomer i midten af proto-porphyrinringen og en femte binding med et proksimalt histidinnitrogen; jern har den sjette frie koordinationsbinding, og det kan binde til ilt.
Når jern er i form af en fri ion, dets type orbitaler d de har alle den samme energi; i myoglobin er jernionen bundet til protoporphyrin og histidin: disse arter forstyrrer magnetisk orbitalerne d noget jern; omfanget af forstyrrelsen vil være forskelligt for de forskellige orbitaler d afhængig af deres rumlige orientering og den forstyrrende art. Da orbitalernes samlede energi skal være konstant, forårsager forstyrrelsen en energisk adskillelse mellem de forskellige orbitaler: energien, der opnås af nogle orbitaler, svarer til den energi, der går tabt af de andre.
Hvis adskillelsen mellem orbitalerne ikke er særlig stor, foretrækkes et elektronisk arrangement med høj centrifugering: bindingselektronerne forsøger at arrangere sig selv i parallelle spins i så mange underniveauer som muligt (maksimal multiplicitet); hvis forstyrrelsen på den anden side er meget stærk, og der er en stor adskillelse mellem orbitalerne, kan det være mere bekvemt at parre bindingselektronerne i orbitalerne med lavere energi (lavt spin).
Når jern binder til ilt, antager molekylet et lavt centrifugeringsarrangement, mens når jern har den sjette koordinationsbinding fri, har molekylet et højt spin -arrangement.
Takket være denne spin -forskel er vi gennem en spektral analyse af myoglobin i stand til at forstå, om ilt (MbO2) er bundet til det eller ej (Mb).
Myoglobin er et typisk muskelprotein (men det findes ikke kun i muskler).
Myoglobin ekstraheres fra kaskelothvalen, hvori den findes i store mængder og renses derefter.
Hvaler har en åndedræt som hos mennesker: med lunger skal de absorbere luft gennem åndedrætsprocessen; kaskelothvalen skal bringe så meget ilt som muligt ind i musklerne, der er i stand til at akkumulere ilt ved at binde den til myoglobinet, der er til stede i dem; iltet frigives derefter langsomt, når hvaleren er nedsænket, fordi dets stofskifte kræver ilt: jo større mængde ilt, som kaskelothvalen er i stand til at absorbere, og jo mere ilt der er tilgængeligt under dykket.
Myoglybin binder ilt på en reversibel måde og er til stede i perifere væv i en større procentdel, jo mere er vævet vant til at arbejde med iltforsyninger, der er fjerne i tid.
<--- Myoglobin er et protein til stede i musklerne, hvis funktion netop er et iltreservoir.

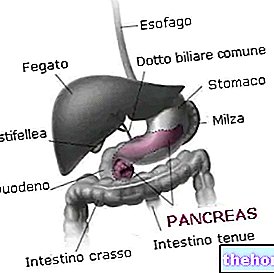
Det, der gør kødet mere eller mindre rødt, er indholdet af hæmoproteiner (det er hæmmen, der gør kødet rødt).
Hæmoglobin har mange strukturelle ligheder med myoglobin og er i stand til at binde molekylært oxygen på en reversibel måde; men mens myoglobin er begrænset til muskler og perifere væv generelt, findes hæmoglobin i erytrocytter eller røde blodlegemer (de er pseudoceller, det vil sige, de er ikke rigtige celler), som udgør 40% af blodet.
I modsætning til myoglobin er hæmoglobins arbejde at tage ilt i lungerne, frigive det i cellerne, hvor det er nødvendigt, tage kuldioxid og frigive det i lungerne, hvor cyklussen begynder igen.
L "hæmoglobin det er et tetrameter, det vil sige, det består af fire polypeptidkæder hver med en hæmgruppe og identiske to og to (i et menneske er der to alfakæder og to betakæder).
Hæmoglobins hovedfunktion er transport af ilt; en anden funktion af blodet, som hæmoglobin er involveret i, er transport af stoffer til vævene.
I vejen fra lungerne (rig på ilt) til vævene transporterer hæmoglobin ilt (samtidig når de andre stoffer til vævene), mens det i den modsatte vej fører det affald indsamlet af vævene, især kulstoffet dioxid produceret i stofskiftet.
I udviklingen af et menneske er der gener, der kun udtrykkes i et bestemt tidsrum; af denne grund er der forskellige hæmoglobiner: føtal, embryonisk, hos den voksne mand.
Kæderne, der udgør disse forskellige hæmoglobiner, har forskellige strukturer, men med nogle ligheder er den funktion, de udfører, mere eller mindre den samme.
En forklaring på tilstedeværelsen af flere forskellige kæder er følgende: i løbet af organismenes evolutionære proces har endda hæmoglobin udviklet sig med speciale i transport af ilt fra områder, der er rige på det, til områder, der er mangelfulde. I begyndelsen af evolutionskæden l "hæmoglobin transporterede ilt i små organismer; i løbet af evolutionen nåede organismerne større dimensioner, derfor blev hæmoglobinet modificeret for at kunne transportere ilt til områder længere væk fra det punkt, hvor det var rig på det; til gør dette, de er i løbet af den evolutionære proces blevet kodet for nye strukturer i kæderne, der udgør hæmoglobin.
Myoglobin binder ilt selv ved beskedne tryk; i perifere væv er der et tryk (PO2) på ca. 30 mmHg: myoglobin ved dette tryk frigiver ikke ilt, så det ville være ineffektivt som iltbærer. Hæmoglobinet derimod , det har en mere elastisk adfærd: det binder ilt til højt tryk og frigiver det, når trykket falder.
Når et protein er funktionelt aktivt, kan det ændre sin form en smule; for eksempel har iltet myoglobin en anden form end ikke-iltet myoglobin, og denne mutation påvirker ikke sine naboer.
Situationen er anderledes i tilfælde af associerede proteiner som hæmoglobin: når en kæde oxygenerer, får den til at ændre form, men denne ændring er tredimensionel, så de andre kæder i tetrameteret påvirkes også. Det faktum, at kæderne er forbundet med hinanden., tyder på, at modifikationen af den ene påvirker de andre naboer, selvom den er i en anden grad; når en kæde oxygenerer, antager tetrameterets andre kæder en "mindre fjendtlig holdning" over for ilt: den vanskelighed, hvormed en kæde kædes oxygenater falder, når kæderne tæt på det oxygenerer igen. Det samme gælder for deoxygenering.
Deoxyhemoglobins kvaternære struktur kaldes T (spændt) form, mens oxyghemoglobins struktur kaldes R (frigivet) form; i den spændte tilstand er der en række temmelig stærke elektrostatiske vekselvirkninger mellem sure aminosyrer og basiske aminosyrer, der fører til en stiv struktur af deoxyhemoglobin (derfor er den "spændte form"), mens når ilt er forbundet, er disse enheders enhed interaktioner falder (derfor den "frigivne form"). Yderligere, i fravær af ilt, stabiliseres ladningen af histidinet (se struktur) af den modsatte ladning af asparaginsyren, mens der i nærvær af ilt er en tendens fra proteinets side til at miste en proton; alt dette indebærer, at iltet hæmoglobin er en stærkere syre end deoxygeneret hæmoglobin: bohr -effekt.
Afhængigt af pH binder hæmgruppen mere eller mindre let til ilt: i et surt miljø frigiver hæmoglobin lettere ilt (den spændte form er stabil), mens bindingen med ilt i et grundlæggende miljø er sværere.

Hvert hæmoglobin frigiver 0,7 protoner pr. Mol ilt (O2), der kommer ind.
Bohr -effekten tillader hæmoglobin at forbedre sin evne til at transportere ilt.
Hæmoglobinet, der bevæger sig fra lungerne til vævene, skal balancere sig selv som en funktion af tryk, pH og temperatur.
Lad os se effekten af temperaturen.
Temperaturen i lungealveolerne er ca. 1-1,5 ° C lavere end den ydre temperatur, mens temperaturen i musklerne er ca. 36,5-37 ° C; efterhånden som temperaturen stiger, falder mætningsfaktoren (ved samme tryk): dette sker, fordi kinetisk energi stiger, og dissociation favoriseres.

Der er andre faktorer, der kan påvirke hæmoglobins evne til at binde til ilt, hvoraf den ene er koncentrationen af 2,3 bisphosphoglycerat.
2,3 bisphosphoglycerat er en metabolisk til stede i erythrocytter i en koncentration på 4-5 mM (i ingen anden del af organismen er den til stede i en så høj koncentration).
Ved fysiologisk pH deprotoneres 2,3 bisphosphoglycerat og har fem negative ladninger på sig; den er klemt mellem de to betakæder af hæmoglobin, fordi disse kæder har en høj koncentration af positive ladninger. De elektrostatiske interaktioner mellem betakæderne og 2,3 bisphosphoglyceratet giver systemet en vis stivhed: der opnås en spændt struktur, der har ringe affinitet til ilt; under oxygenering udvises derefter 2,3 bisphosphoglyceratet.
I erythrocytter er c "et specielt apparat, der omdanner 1,3 bisphosphoglycerat (produceret ved metabolisme) til 2,3 bisphosphoglycerat, så det når en koncentration på 4-5 mM, og derfor er hæmoglobinet i stand til at udveksle" oxygen "i vævene.
Hæmoglobinet, der ankommer til et væv, er i frigivet tilstand (bundet til ilt), men i nærheden af vævet carboxyleres det og passerer til den spændte tilstand: proteinet i denne tilstand har mindre tendens til at binde med ilt, med respekt til frigivet tilstand, derfor frigiver hæmoglobin ilt til vævet; endvidere er der ved reaktion mellem vand og kuldioxid produktion af H + -ioner, derfor yderligere ilt på grund af bohr -effekten.
Kuldioxid diffunderer ind i erythrocyt, der passerer gennem plasmamembranen; da erytrocytter udgør omkring 40% af blodet, bør vi forvente, at kun 40% af kuldioxid, der diffunderer fra vævene, kommer ind i dem, faktisk kommer 90% af kuldioxid ind i erytrocytterne, fordi de indeholder et enzym, der omdanner kuldioxid i kulsyre, resulterer det i, at den stationære koncentration af kuldioxid i erytrocytterne er lav, og derfor er indgangshastigheden høj.

Et andet fænomen, der opstår, når en erytrocyt når et væv, er følgende: ved gradient forlader "HCO3- (derivat af kuldioxid)" erythrocyt, og for at afbalancere output af en negativ ladning har vi "indgang af chlorider, som bestemmer en stigning i osmotisk tryk: for at afbalancere denne variation er der også indtrængen af vand, der forårsager hævelse af erytrocyt (HAMBURGER -effekt) Det modsatte fænomen opstår, når en erytrocyt når lungealveolerne: en deflation af erytrocytterne (HALDANE -effekt) Derfor er de venøse erytrocytter (rettet mod lungerne) rundere end de arterielle.